
A New Genetic Perspective on Gorillas
Categories: Journal no. 42, Eastern gorillas, Western Gorillas, Taxonomy, Gorilla Journal, Gorilla Species
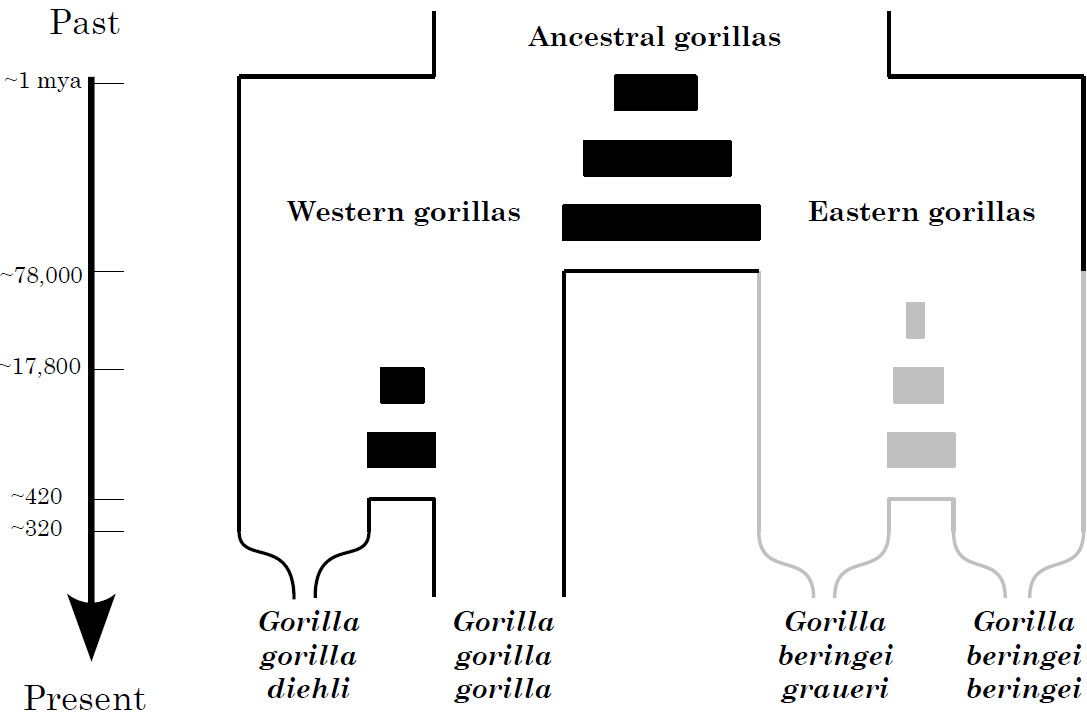
Schematic depiction of the evolutionary histories of gorillas. Times are given in years and represent mode values of parameter distributions which are associated with large confidence intervals. The filled boxes indicate barriers to progressively decreasing gene flow between populations. The gray shading of eastern gorillas should imply that not much is known about the demographic history of these particular gorillas but investigations are underway. (© Olaf Thalmann)
Over the last two decades genetic studies have greatly influenced our understanding of the complex evolutionary history of one of our closest living relatives - the gorilla (Garner & Ryder 1996; Anthony et al. 2007; Thalmann et al. 2007). The contemporary distribution of gorillas in equatorial Africa features a pronounced gap of several hundred kilometers between western (Gorilla gorilla) and eastern gorillas (G. beringei), yet these two species retain behavioural and morphological similarities and were until recently considered subspecies of the same species (Harcourt & Stewart 2007; Groves 1967). Whereas previous genetic studies provided valuable insights into the history of gorillas on an interspecific level (Thalmann et al. 2007; Thalmann et al. 2005), little is known about the intraspecific evolutionary dynamics of gorilla populations.
The focus of our most recent study was a comprehensive assessment of the evolutionary histories of the broadly distributed population of western lowland gorillas (G. g. gorilla) and the range-restricted, critically endangered, Cross River gorillas (G. g. diehli). While some tens of thousands of western lowland gorillas exist, Cross River gorillas number less than 300 individuals found only in highland areas along the Nigeria–Cameroon border (Oates et al. 2007). Previous genetic analyses of these gorillas using multiple autosomal microsatellite loci applied to DNA derived from non-invasively collected fecal samples suggested a recent and/or severe population decrease (Bergl et al. 2008). We built upon this work by generating comparable historical data using DNA obtained from 100-year-old Cross River gorilla museum specimens and comparing the diversity present a century ago and today (Bergl & Vigilant 2007). Our findings suggest that higher genetic diversity was present in the recent past.
We also looked at the deeper demographic history of Cross River gorillas by comparing how well our data supported two different scenarios describing how western lowland gorillas and Cross River gorillas became separate populations. One possibility is that the populations diverged, but at some later point had secondary contact before finally becoming separate once more. Another scenario suggests that the populations diverged but continued to exchange migrants for some time. The theoretical models for each scenario allowed for a severe, recent population decline in the Cross River population and incorporated contemporary and museum samples appropriately. We found that the scenario of secondary contact was not supported, and that the scenario of divergence and continued gene flow was consistent with our data.
These results contribute to a more complete understanding of the evolutionary history of gorillas. An initial split of western and eastern gorillas about one million years ago was accompanied by bidirectional gene flow until approximately 78,000 years ago (Thalmann et al. 2007). Some 17,800 years ago, western lowland and Cross River gorillas diverged but substantial gene flow (~ 4 individuals per generation) between the two western gorilla subspecies ceased as recently as 420 years ago. A marked decline of Cross River gorilla population size began only about a hundred years later. In contrast, the population size of western lowland gorillas increased after the divergence from the Cross River gorillas. These events, along with the signals of likely population structure in the ancestral western gorilla population (Thalmann et al. 2007), are consistent with a scenario of changing climate conditions over the late Pleistocene that led to repeated expansion and contraction of forests, and hence of forest-dwelling ape populations, as well as more recent increased anthropogenic impact. Hunting of gorillas continues today and represents one of the greatest threats to their survival (Oates et al. 2007). Particularly for small populations that may have gone through recent reductions in size, effective enforcement of anti-poaching laws and maintenance of existing habitat to allow the population to stabilize and expand is crucial to their future survival.
Olaf Thalmann, Richard A. Bergl and Linda Vigilant
Original publication:
Thalmann, O., Wegmann, D., Spitzner, M., Arandjelovic, M., Guschanski, K., Leuenberger, C., Bergl, R. A. & Vigilant, L. (2011): Historical sampling reveals dramatic demographic changes in western gorilla populations. BMC Evolutionary Biology 11, 85
References
Anthony, N. M. et al. (2007): The role of Pleistocene refugia and rivers in shaping gorilla genetic diversity in central Africa. Proc. Natl. Acad. Sci. U. S. A. 104 (51), 20432-20436
Bergl, R. A. & Vigilant, L. (2007): Genetic analysis reveals population structure and recent migration within the highly fragmented range of the Cross River gorilla (Gorilla gorilla diehli). Mol. Ecol. 16 (3), 501-516
Bergl, R. A. et al. (2008): Effects of habitat fragmentation, population size and demographic history on genetic diversity: The Cross River gorilla in a comparative context. Am. J. Primatol. 70 (9), 848-859
Garner, K. J. & Ryder, O. A. (1996): Mitochondrial DNA diversity in gorillas. Mol. Phylogenet. Evol. 6 (1), 39-48
Groves, C. P. (1967): Ecology and taxonomy of the gorilla. Nature 213 (5079), 890-893
Harcourt, A. H. & Stewart, K. J. (2007): Gorilla society: What we know and don't know. Evolutionary Anthropology: Issues, News, and Reviews 16 (4), 147-158
Oates, J. et al. (2007): Regional action plan for the conservation of the cross river gorilla (Gorilla gorilla diehli) (IUCN/SSC Primate Specialist Group and Conservation International, Arlington, VA, USA)
Thalmann, O. et al. (2007): The complex evolutionary history of gorillas: Insights from genomic data. Mol. Biol. Evol. 24 (1), 146-158
Thalmann, O. et al. (2005): Nuclear insertions help and hinder inference of the evolutionary history of gorilla mtDNA. Mol. Ecol. 14 (1), 179-188